
Learn how Cyagen’s PD-L1 conditional knockout (cKO) mice have been used to advance immunity and intestinal microbiology research
The intestinal tract plays an overwhelmingly significant role for digestion and nutrient absorption, but it is also the largest immune organ. Homeostasis of the intestinal epithelium is maintained through the intestinal microbiome, immune cells, and mucosal barrier, which together act as one of the most important safeguards for human health.
The article entitled “Niche-specific MHC II and PD-LI regulate CD4+CD8aa+ intraepithelial lymphocyte differentiation,” published in the Journal of Experimental Medicine, first revealed the molecular mechanisms of intestinal epithelial cells (IECs) regulating T cell differentiation (IEL, intraepithelial lymphocytes). In addition, findings indicate the microbiome essential for the epithelial differentiation of MHCII, PD-L1, and IEL in the small intestine.

Conventional CD4+ T cells are differentiated into CD4+CD8αα+ intraepithelial lymphocytes (IELs) in the intestine; however, the roles of surrounding intestinal epithelial cells (IECs) are poorly understood. According to findings published in the Journal of Experimental Medicine [1] by the Korean team led by Dr. Moon, in the distal small intestine, IECs, stimulated by microbiota and IFN-γ, can provide niche adaptive signals to promote the differentiation of CD4+CD8αα+IELs (DP [double positive] IELs) by expressing MHC class II molecules and programmed death ligands-1 (PD-L1).
Notes: All mice herein are of C57BL/6 genetic background. SP refers to CD4+ single positive cells; DP refers to CD4+CD8+ double positive cells.
In order to explore the IECs’ atypical APC role during DP IELS differentiation, researchers analyzed the expression of MHC II on IECs in different sites of the small intestine of C57BL/6 wild-type mice. A series of flow cytometry experiments and RNA-seq experiments have shown that the role of IECs in the differentiation of DP IELS is recursive related to MHC II antigen presentation. In addition, to identify the role of MHC II expressed by IECs in the differentiation of DP IELs, researchers used mice with specific knockout of MHC II in IECs (MHC II△IEC) to confirm that the differentiation of IELs requires expression of MHC II on IECs, as compared with the MHC IIfl/fl control mice (Figure 1G-H).
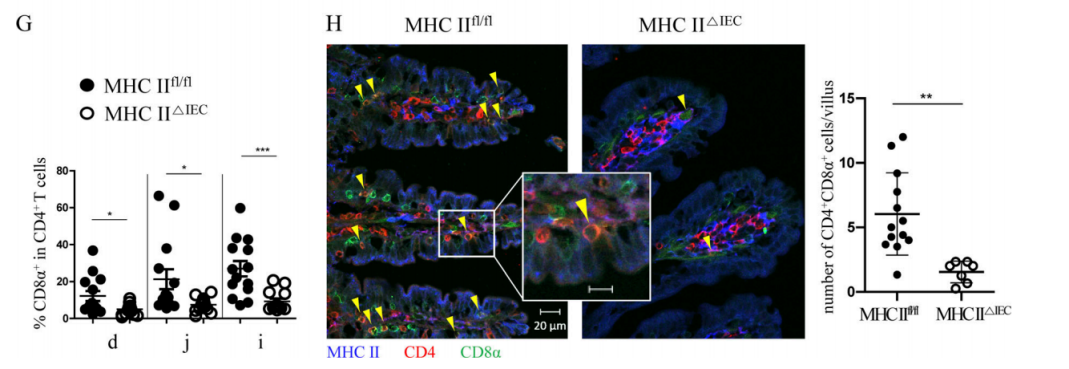
Figure 1G-H. Both flow cytometry (G) and immunofluorescence experiments (H) showed that the proportion of DP IELs in SP IELs cells decreased significantly in MHC II△IEC mice, compared with the MHC IIfl/fl mice. Immunofluorescence experiments (H) also showed that the majority of DP IELs contacts the base lateral surface of epithelial cells with highly expressed MHC II.
It has been reported that IFN-γ can strongly induce the expression of MHC II in non-hematopoietic cells (such as IECs). To further verify this claim, researchers used a kind of IFN-γ receptor knockout mice (IFN-γR−/−). The research findings echo with the previous claim: IECs of IFN-γR−/−mice did not have the expression of MHC II detected, the differentiation of DP IELs was severely blocked, and its proportion in SP T cells was almost 0 (Figure 2A). In wild-type mice, after adding anti-IFN-γ antibody, the expression level of MHC II and the proportion of DP in SP IELs cells were significantly reduced (Figure 2B). Other reports also show that IFN-γ is vital for the differentiation of DP IELs, which can promote the differentiation of DP IELs by inducing the expression of the transcription factor T-bet. It means that IFN-γ plays an important role in the functional maturation of DP IELs.
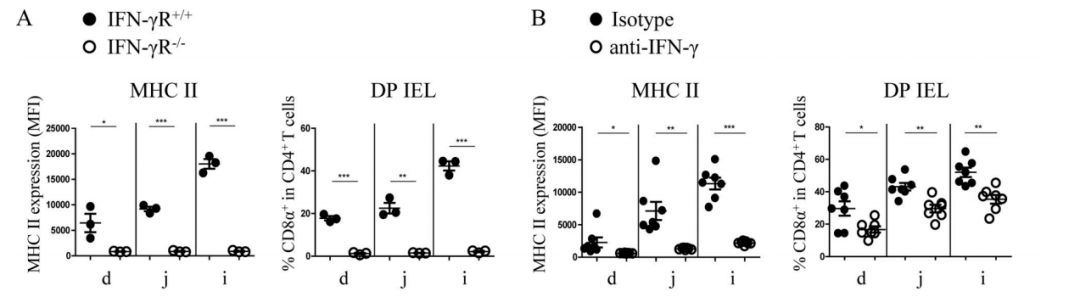
Figure 2. IFN-γ can strongly induce the expression of MHC II in non-hematopoietic cells (such as IECs).
Bone marrow (BM) chimera mice were developed by researchers to explore the effect of IFN-γ on the differentiation of DP IELs in IELs and IECs. BM cells were obtained from the leg bones of donor mice and injected intravenously into the recipient mice irradiated with a lethal dose (hematopoietic cells of donor mice→recipient mice). After 8 weeks of hematopoietic reconstitution period, the recipient mice were sacrificed. The results (Figure 2C) showed that the complete IFN-γR signals are indispensable for differentiation of DP IELs, whether in hematopoietic cells (e.g.CD4+IELs) or non-hematopoietic cells (e.g. IECs). Significant differences in the expression quantity of MHC II and the proportion of DP IELS as observed in different areas of small intestine of wild-type mice were not detected in BM chimeric mice. Researchers theorized that the recipient mice after immune reconstitution might produce more IFN-γ than mice in steady-state, which inflamed the intestinal microenvironment and thus eliminated the regional difference.
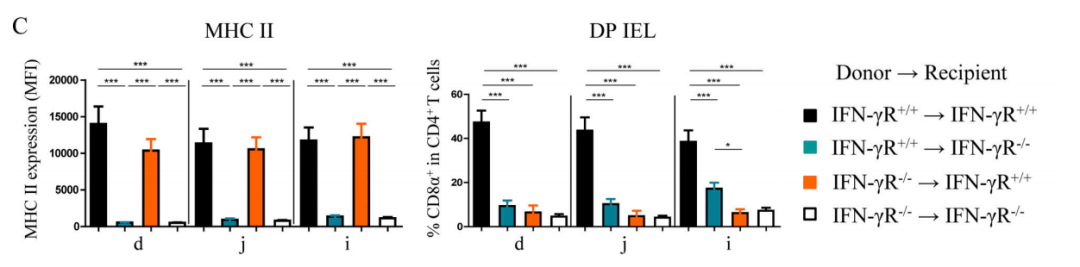
Figure 2C. Complete IFN-γR signals are indispensable for differentiation of DP IELs.
In order to directly explore whether MHC II+IECs play an atypical role in the differentiation of DP IELs, researchers constructed small intestinal organoids derived from IFN-γR+/+ mice and IFN-γR−/−mice. They first verified that IFN-γ can exert the same effect in organoids as in vivo - IFN-γ can strongly induce the expression of MHC II on IECs in organoids (Figure 2D-E). Besides, other research findings also showed that T-box expressed in T cells (T-bet) is an important upstream regulator during DP IELs differentiation, and it can induce the expression of Runx3 and inhibit the expression of ThPOK. With the help of intestinal microenvironmental stimulators, the cytokines, such as IFN-γ induced by T-bet, can further promote the differentiation of DP IELs. Therefore, researchers co-cultured the small intestinal organoids pre-stimulated by IFN-γ and ovalbumin peptide (OVAp) and OVA-specific CD4+T cells (OT-II) pre-stimulated by anti-CD3ε/CD28 for 24 hours in the presence of intestinal microenvironmental stimulators TGF-β and retinoic acid (RA) (Figure 2F). The results showed that IFN-γ could induce a large amount of expression of MHC II, and MHC II+IECs played an atypical APC role in the DP IELs differentiation, and TGF-β and RA stimulation could further promote the differentiation of DP IELs.
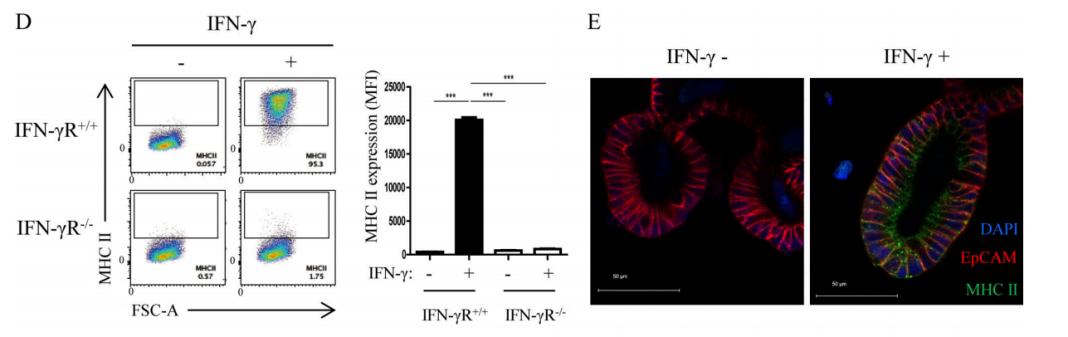
Figure 2D-E. Flow cytometry (D) and immunofluorescence experiments (E) showed that after IFN-γ stimulation, the expression level of MHC II on the organoids IECs derived from IFN-γR+/+ mice increased significantly, while MHC II expression was not available on the organoids IECs derived from IFN-γR−/−mice.
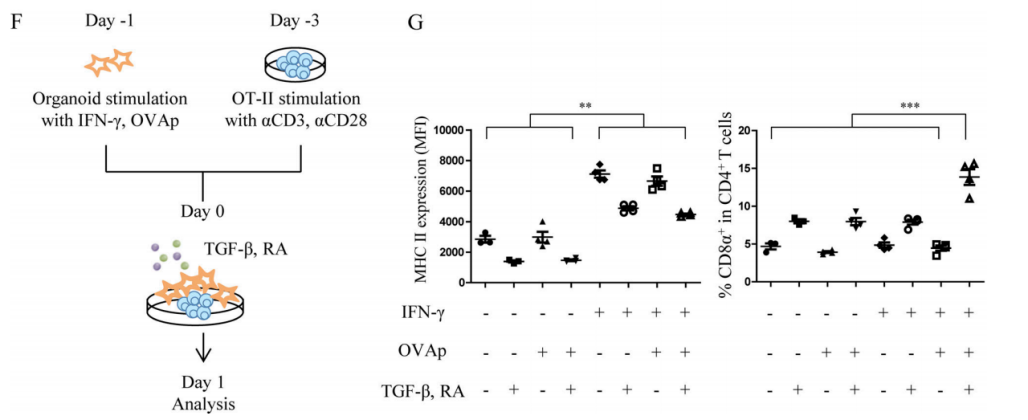
Figure 2F-G. Co-culture experiment (F); MHC II expression on IECs (G left); DP ratio in SP OT-II cells (G right).
The experiments above show that MHC II+IECs may act as APC for homologous stimulation of CD4+IELs. Researchers thus speculated that they could also express other co-receptors that coordinate with MHC II to regulate T cells. Since MHC II expression on IECs depends on IFN-γ, researchers further speculated that these co-receptors should be related to the IFN-γ signal in IECs. Through the RNA-seq analysis of small intestinal organoids with/without IFN-γ treatment, researchers revealed that the Cd274 gene coding PD-L1 protein was significantly correlated with the expression of MHC II-related genes (Fig. 3A-B). Both organoid experiment (Fig. 3C) and in vivo experiment (Fig. 3D-E) indicated that IFN - γ could significantly affect the expression quantity of PD-L1. To further verify that differentiation of DP IELs requires expression of PD-L1 on IECs, the researchers used PD-L1 knockout mice (PD-L1−/−), RAG-1 knockout mice (RAG-1−/−, mice without mature T cell and B cell), mice with specific knockout of PD-L1 target gene in IECs (PD-L1△IEC), and PD-L1fl/fl control mice constructed by Cyagen. Results drawn from these experiments (Fig. F-H) show that PD-L1 on IECs plays an important role in the differentiation of DP IELs.
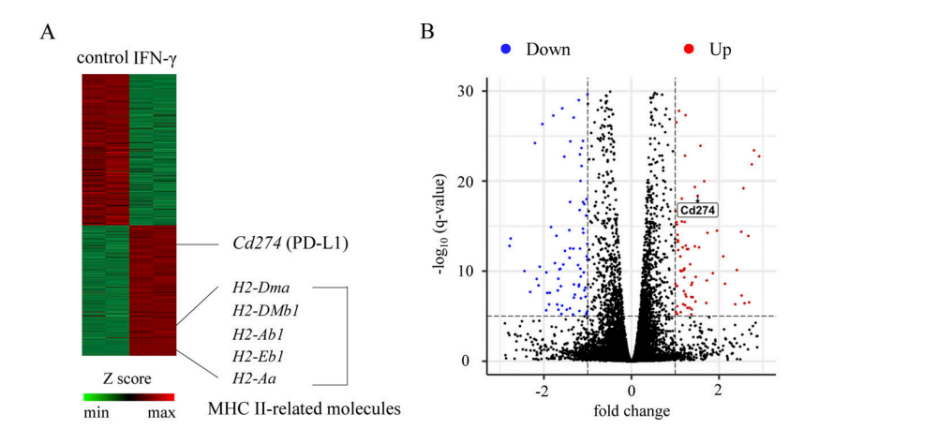
Figure 3A-B. Heatmap of differentially expressed genes (A) and volcano plot (B).
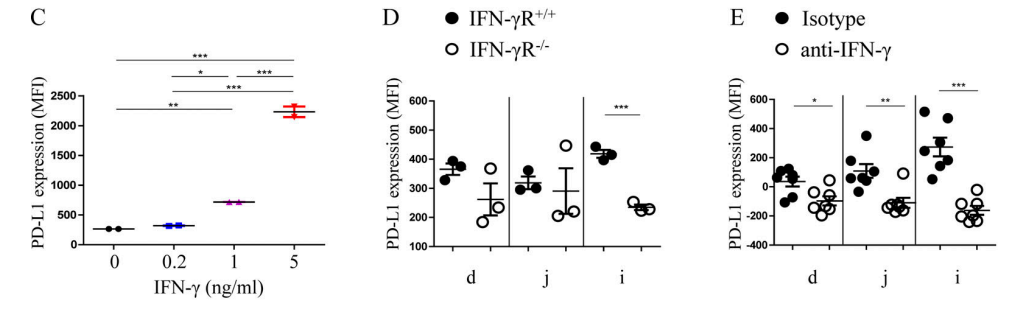
Figure 3C-E. (C) The expression quantity of PD-L1 in small intestinal organoids after IFN-γ treatment at varying concentrations; (D) The expression quantity of PD-L1 in IFN-γR−/−mice decreased significantly compared with IFN-γR+/+mice; (E)The expression quantity of PD-L1 on IECs decreased significantly after injection of anti-IFN-γ antibody in IFN-γR+/+mice.

Figure 3F-H. (F) Compared with PD-L1+/+mice, the proportion of DP IELS decreased significantly in the ileum of PD-L1−/−mice; (G) After RAG-1−/− mice were injected with splenic CD4+ T cells, and DP IELs development were inhibited by anti–PD-1 treatment; (H) Compared with PD-L1fl/fl mice, the proportion of DP IELS decreased significantly in the ileum of PD-L1△IEC mice.
It has been reported that both the production of IFN-γ in the small intestine and the expression of MHC II on IECs require the presence of microbiota. Therefore, to explore whether the expression of PD-L1 on IECs is regulated by the microbiota, germ-free (GF) mice and specific pathogen-free (SPF) mice were used for experiments. According to the experimental findings, microbiota-induced IFN-γ can stimulate expression of MHC II and PD-L1 on IECs, both of which may function to regulate DP IELs differentiation.
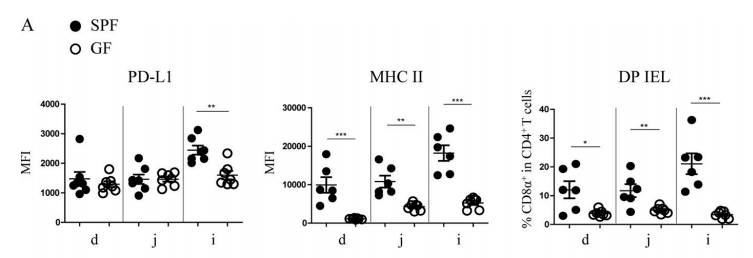
Figure 4A. Compared with SPF mice, the expression level of PD-L1 on IECs of GF mice decreased significantly in ileum, while the expression quantity of MHC II and the proportion of DP IELs decreased significantly in various areas of small intestine, especially in ileum.
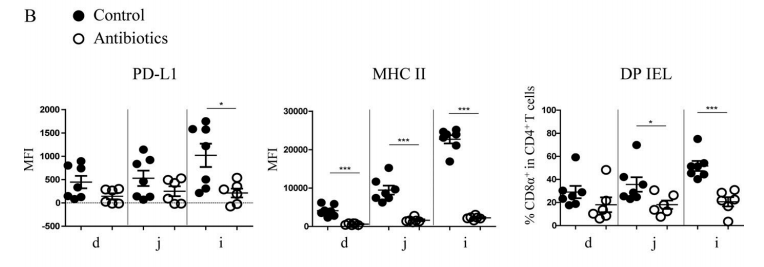
Figure 4B. After antibiotic treatment, the expression level of PD-L1 on IECs of SPF mice decreased significantly in ileum, and the expression quantity of MHC II and the proportion of DP IELs decreased significantly in various areas of small intestine, especially in ileum.
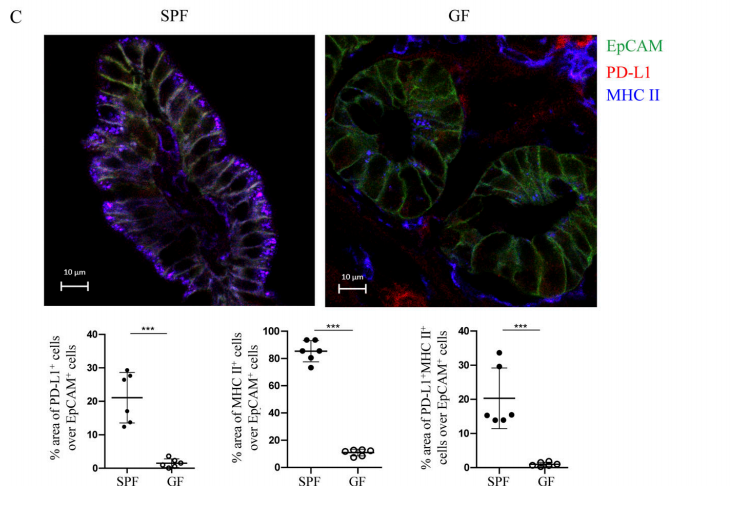
Figure 4C. According to immunofluorescence experiments, most IECs in the ileum of SPF mice expressed MHC II, and 20% expressed PD-L1, while in GF mice, the expression of MHC II and PD-L1 were significantly reduced.
Conventional CD4+ T cells and CD8+ T cells maintain their own lineages by expressing T helper-inducing POZ/Krüppel-like factor (ThPOK) and runt-related transcription factor 3 (Runx3), respectively. Therefore, the differentiation of DP IELs requires the up-regulation of Runx3 expression and the deletion of ThPOK. In addition, the expression of MHC II and PD-L1 on IECs are critical to the differentiation of DP IELs, which indicates that the signals released by TCR and PD-1 may participate in the cell reprogramming of CD4+ IECs. Previous experimental results showed that endogenous PD-1 signal of T cells was essential to the differentiation of DP IELs (Fig. 5A-C). To further explore the molecular mechanism involved, ThPOK-GFP reporter mice were used. The experimental results show that the PD-1 inhibition of ThPOK expression and the differentiation of DP IELs require the presence of PD-1 signals (Figure 5D-E). In addition, the in vitro experimental results (Figure 5G-I) showed that PD-1 signal down-regulates the expression of ThPOK in CD4+IECs through the classical Src homology 2 domain–containing tyrosine phosphatase (SHP) pathway, thereby promoting the differentiation of DP IELs.
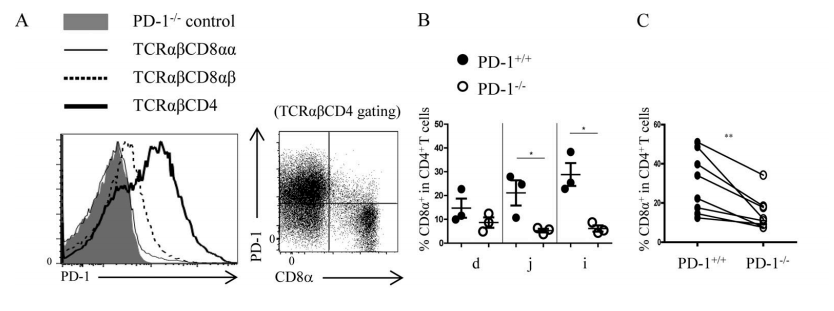
Figure 5A-C. (A) The expression of PD-1 in different IEL subgroups in C57BL/6 wild-type mice. When SP IELs differentiated into DP IELs, the PD-1 expression decreased. (B) Compared with PD-L1+/+ mice, the proportion of DP IELs decreased significantly in the small intestines of PD-L1−/− mice. (C) After 1:1 mixing splenic CD4+T cells from PD-L1+/+ and PD-L1−/− mice, they were transplanted into RAG-1−/− recipient mice. The proportion of DP IELs from PD-L1−/− CD4+ T cells decreased significantly.
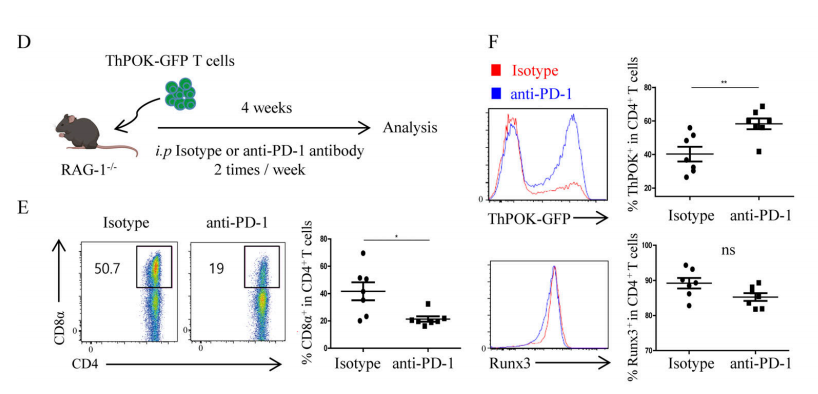
Figure 5D-F. (D) Experimental diagram. Splenic T cells from ThPOK-GFP reporter mice were transplanted into RAG-1−/−recipient mice, and anti-PD-1 antibody treatment was carried out on the recipient mice during the reconstruction period, (E) The expression levels of ThPOK and Runx3; (f) The proportion of DP IELs.
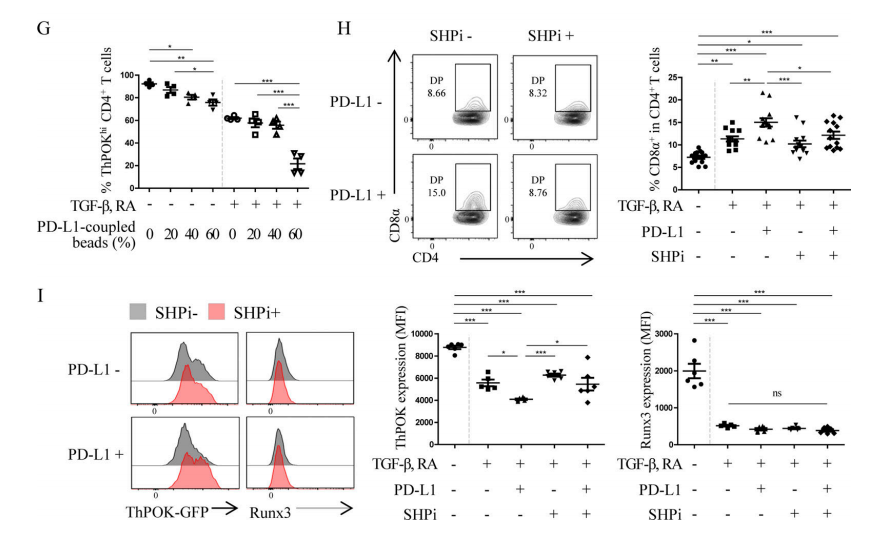
Figure 5 G-I. (G) After in vitro co-culture of CD4+T cells, TGF-β, RA, and PD-L1 for 3 days, the proportion of ThPOKhi cells was determined. PD-L1 mediated the down-regulation of ThPOK expression. (I-H) After in vitro co-culture of CD4+T cells, TGF-β, RA, PD-L1 and SHP inhibitor (SHPi) for 3 days, the proportion of DP IELs and the expression levels of ThPOK and Runx3 were determined.
In summary, this paper has demonstrated that IECs are an important regulator to promote differentiation of intestinal DP IELs and play a role as atypical APC. IFN-γ produced by intestinal microbiota can stimulate expression of MHC II and PD-L1 on IECs, and provide TCR stimulation and co-receptor signal to SP IELs, thereby promoting differentiation towards DP IELs. In addition, researchers found that the unique gene expression profiles of IECs at different physiological locations may also affect tissue-resident immune cells around them (such as differentiation of DP IELs).
Note: All mice herein are the genetic background of C57BL/6
As the second part of deciphering the human genome, the human gut microbial genome acts as the immediate indicator of a person’s health. Large quantities of research have proven that intestinal microorganisms are closely related to the occurrence and development of cardiovascular, immune, nervous, and digestive diseases, as well as tumors - the underlining causal relationship has gradually become clearer. Cyagen's all-in-one innovative CRO service network platform can provide genetically engineered mouse models, GF mouse models, breeding, and downstream model evaluation services to meet the requirements of any experimental animal model project.
Large quantities of research proved that intestinal microorganisms are closely related to the occurrence and development of cardiovascular, immune, nervous, digestive diseases as well as tumors. Cyagen's all-in-one innovative CRO service network platform can provide genetically engineered mouse models, Ready-to-use models, drug development animal modelsand model supporting services for your demands on experimental animal models:
Custom Genetically Engineered Mouse and Rat Models
Cyagen Ready-to-use Models
The Cyagen AI Knockout Mouse Model eBank provides a searchable listing of over 16,000 custom KO and cKO/floxed strains from which live mice are available to researchers in as few as 2 weeks.
Drug Development Animal Models
Professional Model Supporting Services
In additional model construction,. we also offer phenotypic characterization and other downstream services required to establish experimental models for immunity and intestinal microbiology studies.
Contact us with your project details to request your complimentary model generation project consultation, strategy, and quote.
Reference:
Moon S , Park Y , Hyeon S , et al. Niche-specific MHC II and PD-L1 regulate CD4+CD8αα+ intraepithelial lymphocyte differentiation[J].
Journal of Experimental Medicine, 2021, 218 (4).DOI: 10.1084/jem.20201665
We will respond to you in 1-2 business days.